 Pigmy Hedgehog Tenrec Echinops telfairi at 1 day old.
Pigmy Hedgehog Tenrec Echinops telfairi at 1 day old.
Click on the photo or here to view a larger version.
Back to Tenrec Resources and Information
The reproduction and development of the Pigmy Hedgehog Tenrec Echinops telfairi
By Gillian K. Godfrey, M.A., D.Phil. and William Oliver, MSc. Research Assistant Jersey Wildlife Preservation Trust.
Resumé
A colony of the Pigmy Hedgehog Tenrec Echinops telfairi has been maintained at the Trust since the original acquisition of 2 males and 2 females from the Amsterdam Zoo in December 1966. The stock has flourished since that time and has bred successfuly every year, though consistent inbreeding and a poor sex ratio led to a further importation of 1.1 in April 1975 and a further male in October of the same year from a private collector in West Germany. During the twelve year period to 1978 at least 135 Echinops have been born from a total of 32 parturitions. Of these, 97 Echinops have been successfully reared and 50 specimens have been exported to other collections in Europe and the U.S.A.
The early history and management of this species has been variously described by Ormrod (1967), Mallinson (1968), Scaling (1971) and Riordan (1972), and the purpose of this paper is to present an analysis of their breeding record, together with some observations on their reproduction and ontogeny which have not been described before. Nonetheless, it is important to note that a considerable literature on the reproduction and behaviour of Echinops already exists, notably by Herter (1962, 1963), Honegger and Noth (1965), Eisenberg and Gould (1966, 1970), Eisenberg and Muckenhirn (1968) and Eisenberg (1976). In order to put our own observations into perspective, it has been necessary to summarise some of this information, though for a detailed account of certain aspects, e.g. social and reproductive behaviour and endogenous rhythms, the original papers should be consulted.
Similarly, the practical management of the animals at the Trust has been extensively covered by the earlier authors in the Annual Reports. It is unnecessary to reiterate much of this information, except to mention a single important change in husbandry: the original 'Tenrec mixture and Complan supplement' described by Mallinson has now been discontinued since the mixture was relatively unpalatable and deterioriated rapidly. The animals tended to subsist largely on the Complan element of the diet which led to excessive weight gains and plague deposition on the teeth. This mixture has, therefore, been replaced by a basic diet of whole minced chicks (eviscerated) to which raw beef heart, liver or kidney has been added in the ration of 4:1 fresh weight. The new mixture is evidently higher palatable and has a better roughage content. Soft fruit and vitamin/mineral supplementation remains the same as that previously described, as does the twice weekly substitution of this mixture by offering mealworms, crickets and/or pink mice.
The Annual Cycle
It is well known that Echinops exhibit a marked annual cycle in activity which corresponds to seasonal variation in their native habitat in south-western Madagascar. A relatively long period of sustained inactivity and low metabolic rate (hibernation) which lasts from 3-5 months and corresponds with the austral winter, is succeeded by a longer period of normal activity and reproduction which corresponds to the austral spring and summer. This alternation in activity levels is undoubtedly adaptive since torpor is maintained over the period when conditions are least favourable, i.e. relatively low temperatures, high aridity and a scarcity of small animal prey. In the laboratory, the mainpulation of tenperature gradients and the deprivation of food, have both been found to induce or prolong torpor (Eisenberg & Gould 1970). Humidity levels may also be significant, but no evidence has been put forward that variation in day length is a contributing factor.
Pigmy Hedgehog Tenrec Echinops telfairi at 1 day old.
Click on the photo or here to view a larger version.
However, although within any given population the synchronisation of seasonal periodicity is affected by ambient temperature and the availability of food, the influence of these conditions is by no means absolute. Indeed, individual animals maintained under similar conditions appear to show considerable variation in the propensity to become torpid. Thus, certain individuals have occasionally become torpid without any external manipulation of cage temperature or contrarily, spontaneous temporary re-emergence from torpor has occurred in one or more individuals while others remained torpid. Similarly, it has been noted that the entry into, or arousal from torpor, by some of the individuals within a captive population has also occurred prior to the adjustment of temperature and humidity. These variations may, in part, be attributable to an endogenous rhythm carried over in newly imported animals unsynchronised to the imposed annual regimen. Wild caught animals, for instance, have been known to maintain their annual rhythm with respect to a southern hemisphere pattern for at least the first year after importation to the United States (Eisenberg & Muckenhirn, 1968) and comparable observations have been made here with stock acquired from other collections.
It is probable that inconsistencies in torpidity amongst our own colony however, may also reflect a failure to maintain optimal seasonal temperature/humidity gradients since cage design is at fault and precise manipulation of ambient conditions is not possible. The precise control of the cage environment is certainly desirable for the general maintenance of tenrecs, and Herter (1962) has demonstrated the correlation between ambient and cloacal temperatures as part of an endogenous circadian rhythm in metabolic rate.
In the colony maintained at the Trust, hibernation approximates to a normal northern hemisphere schedule and usually lasts from late November until late March. Towards the end of the active phase, maximum weights are attained by increased appetite and rapid fat deposition (Fig. 1). The onset of hibernation is encouraged by the lowering of cage temperatures from 27-30°C to 21-24°C, a gradient similar to average values derived from field measurements (Eisenberg & Gould, 1967).
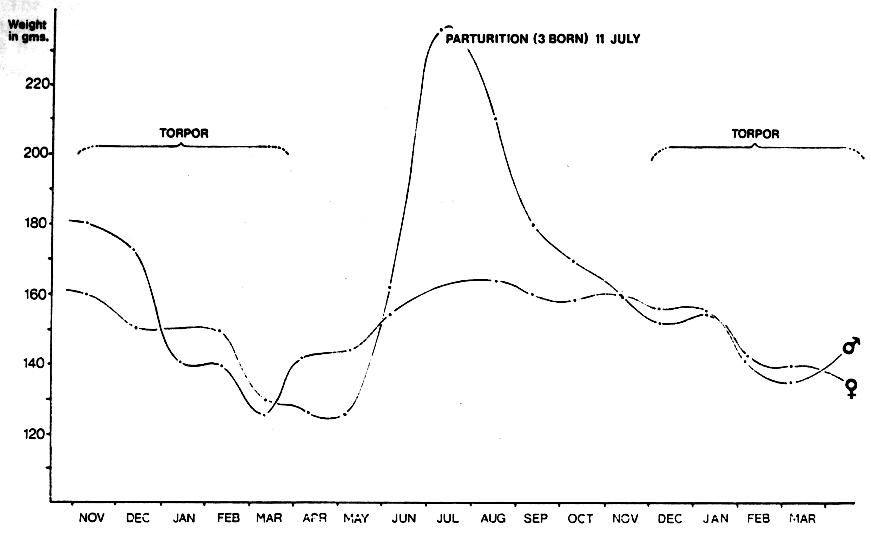 Fig. 1 Annual activity cycle of Echinops telfairi illustrated by the seasonal weight changes of a breeding pair.
Fig. 1 Annual activity cycle of Echinops telfairi illustrated by the seasonal weight changes of a breeding pair.
Click on the figure or here to view a larger version.
Increasing inactivity and loss of appetite follows with a corresponding decline in metabolic rate and rectal temperatures approaching the ambient. The animals are disturbed as little as possible, though fresh water is always made available. In view of the variation in individual torpidity already outlined, food is offered once or twice weekly, although it is frequently left untouched by all individuals.
Emergence from torpor is similarly encouraged by the raising of cage temperature and the liberal provision of preferred foodstuffs such as live insects. Humidity is also increased by the use of a sprinkler two or three times weekly (sprinklers are not used during the torpor period). Animals emerging from torpor rapidly regain normal weight and assume reproductive condition within 2-3 weeks. No difficulty has ever been experienced in stimulating re-emergence from torpor in Echinops.
The Oestrous Cycle
Method: As far as is known, the oestrous cycle in Echinops telfairi has never been investigated, and in 1978 a preliminary study of this aspect of their reproductive biology was begun, using the technique of vaginal (i.e. cloacal) smearing. It has long been known that the squamous cells lining the wall of the vagina become cornified under the influence of oestrogens, and that when the oestrogen-level suddenly falls at the time of ovulation, these cells slough off and pass into the vaginal lumen. Since cornified cells are normally only found in the smears in high numbers during the period of oestrous, their presence provides a useful indicator of this event.
 Pigmy Hedgehog Tenrec Echinops telfairi at 4 day old. Note variation in size, colour and degree of spinescence.
Pigmy Hedgehog Tenrec Echinops telfairi at 4 day old. Note variation in size, colour and degree of spinescence.
Click on the photo or here to view a larger version.
We started smearing soon after the females arouse from torpor in early May, and finished a few days before the litters were born in July. Three females were smeared at weekly intervals during this period, while two further females (M547a and b) were smeared daily for the first 30 days and weekly thereafter. Females were kept in pairs, but because of the shortage of cages the male was present continuously throughout oestrous and pregnancy, except in one female (M547a) where the male was removed when oestrous was at its height and returned 3 weeks later.
Smears were taken with a Pasteur pipette and normal saline solution and inspected while fresh under x 10 magnification for motile sperm; they were fixed while wet with polyethylene glycol and stained by a Papanicolaou method (Baker, Silverton & Luckcock, 1966). Three different types of epithelial cell in the stained smears were identified under x 40 magnification, following a system of classification used in human cytological studies (Wied & Bibbo, 1971). We shall be mainly concerned here with the cornified superficials or 'typical squames'.
In each of the stained smears, relative abundance of cornified cells was estimated by counting the numbers present in a series of Low Power fields; the mean value, when expressed as a percentage of all mature cells (non-cornified as well as cornified), gives the karyopycnotic index or KPI. Total cell-count, which includes all types of squamous cell, was scored subjectively on a 0-5 scale of abundance. Polymorphonuclear leucocytes were scored in a similar manner.
Results: Cornified cells were found to be present in the smears of female 547b in relatively high numbers for at least 19 consecutive days (Fig. 2).
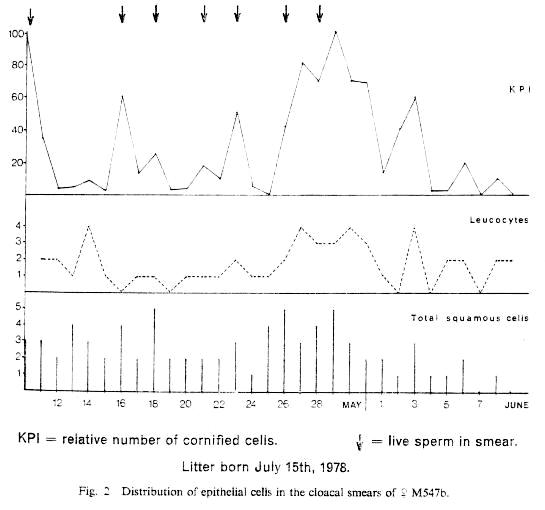 Fig. 2 Distribution of epithelial cells in the cloacal smears of female M547b.
Fig. 2 Distribution of epithelial cells in the cloacal smears of female M547b.
Click on the figure or here to view a larger version.
However, KPI's show five distinct peaks and there is an obvious periodicity, with a mean interval between peaks of 6.0 days. Leucocyte peaks which are less marked are seen on the same day or one day after the peaks in cornification. Seven copulations were recorded during the 19 days and there is clearly a high correlation between the occurrence of copulation and peak cornification.
During the first half of June, the smear-records of female 547a also showed prominent and fairly regular peaks in cornification, with intervals between peaks of 4, 6 and 13 days respectively (Fig. 3).
 Fig. 3 Distribution of epithelial cells in the smears of female M547a.
Fig. 3 Distribution of epithelial cells in the smears of female M547a.
Click on the figure or here to view a larger version.
During the 23 days in May when the female was separated from the male, KPI's showed only one peak in cornification, soon after the male was removed; however, when the total cell-counts for this period were plotted, peaks are evident and the periodicity is seen to be maintained (Fig. 3, lower graph). Seven copulations were recorded.
Smears taken from five females at regular intervals during pregnancy showed a progressive decrease in cell-maturity and cornification as pregnancy advanced. Individual smears occasionally revealed unusual features: for example, a smear from female 500b showed a massive infiltration of RBC's 23 days before parturition. The relatively small size of this litter (4) suggests that embryonic resorption may have been taking place. (In the white-toothed shrew, Crocidura suaveolens, which is being studied at the Trust, high numbers of RBC's in the smears of pregnant females have been found to be associated with foetal mortality, Godfrey, in prep). Further evidence of foetal mortality in Echinops was indicated when a female's body-weight dropped suddenly about 4 weeks before parturition. Only 4 young were born.
The gestation period in Echinops telfairi has been estimated to lie between 62-65 days (Eisenberg & Muckenhirn, 1968) and 61-64 days (Mallinson, 1972). These figures are based on intervals between known copulations and birth. Although data obtained in 1978 from 4 out of 5 females do not confilct with these estimates, a shorter period seems likely.
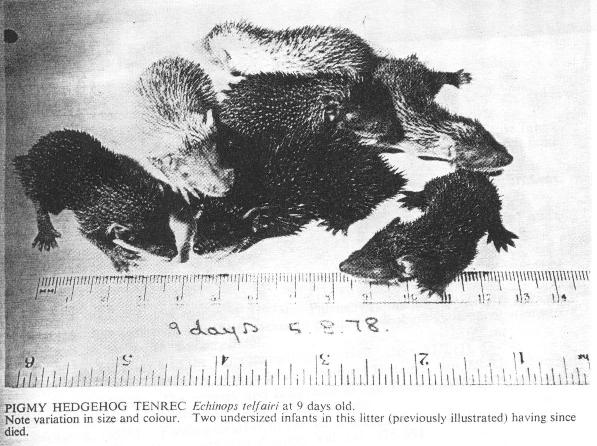 Pigmy Hedgehog Tenrec Echinops telfairi at 9 days old. Note variation in size and colour. Two undersized infants in this litter (previously illustrated) having since died.
Pigmy Hedgehog Tenrec Echinops telfairi at 9 days old. Note variation in size and colour. Two undersized infants in this litter (previously illustrated) having since died.
Click on the photo or here to view a larger version.
Thus in female 547b, (Fig. 2) the interval between the first recorded copulation and birth was 67 days; intervals between the next 5 copulations and birth were 61, 59, 56, 54 and 51 days, while between the last copulation, when the peak in KPI's was the largest, and birth the interval was only 49 days, (When there are several copulations in succession, the last one is usually the most likely to be effective).
With female 547a, the male was removed for 23 days during the middle of the breeding period. The interval between the last copulation before the male was removed (May 9) and birth was 78 days (which is clearly too long), while the interval between the first copulation after the male was returned (June 3) and birth was 54 days (Fig. 3). However, there were two further peaks in cornification after this date, each accompanied by a copulation, and the intervals between these copulations and birth were 49 and 42 days respectively. In the light of these findings a gestation period in excess of 60 days now seems unlikely, and in fact the present data provide strong presumptive evidence of an interval between copulation and birth of under 50 days.
Under the conditions of husbandry described here, Echinops is seen to have a breeding season which begins soon after the females arouse from torpor and lasts for 3 or 4 weeks. The regular changes in cell-type observed over this period suggest that there may be a succession of oestrous cycles, each lasting about 6 days. But whether the first few cycles are sterile as they are in many seasonally breeding mammals cannot, of course, be determined without examining the ovaries.
An interesting feature of this work has been the finding that the cells lining the walls of the vagina do not become cornified if the male is absent, presumably because the oestrogen levels are too low. Smears taken at this time consist mainly of epithelials which are either non-cornified or degenerating, typical squames being rarely found.
Parturition and Parental Care
In view of the variations in seasonal torpor already described, it is not surprising that the 32 parturitions recorded here between 1967 and 1978 have been distributed over the relatively long period from late June to mid-September. However these parturitions follow an approximately normal distribution with a main peak corresponding to a period some 12-14 weeks after emergence from torpor and 9-11 weeks after the commencement of breeding activity (see Fig. 4). Moreover, most of the disparity caused by late season parturitions may be explained in terms of late pairings and/or new importations. Litter sizes have ranged from 1-9 with a mode of 5 (n=25 litters of known size). This compares with a range of 1-10 with a mode of 7 (n=13) parturitions recorded by Eisenberg and Muckenhirn (1968). Curiously enough, a litter of 7 has never been recorded here (Fig. 5).
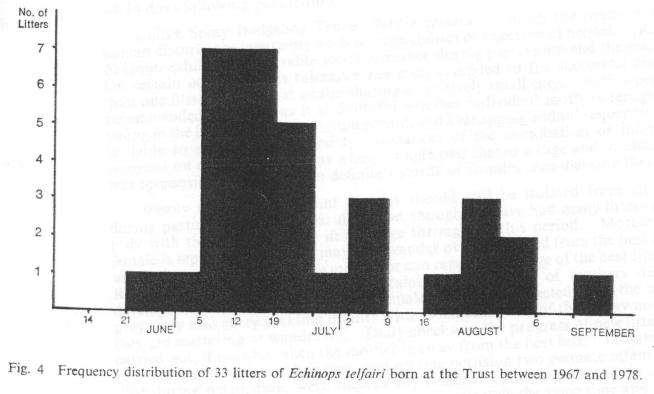 Fig. 4 Frequency distribution of 33 litters of Echinops telfairi born at the Trust between 1967 and 1978.
Fig. 4 Frequency distribution of 33 litters of Echinops telfairi born at the Trust between 1967 and 1978.
Click on the figure or here to view a larger version.
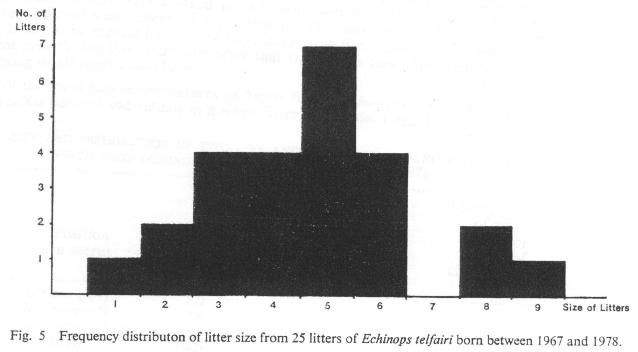 Fig. 5 Frequency distribution of litter size from 25 litters of Echinops telfairi born between 1967 and 1978.
Fig. 5 Frequency distribution of litter size from 25 litters of Echinops telfairi born between 1967 and 1978.
Click on the figure or here to view a larger version.
Parturition always takes place in the next box and pre-natal females indulge in considerable nest building activity. Late-term animals are given leaves, grass, paper, woodwool, hay etc. for this purpose. Nipple enlargement occurs about four weeks before birth; perineal swelling has been recorded up to 5 days pre-partum. Two or more nest boxes are also provided, since nest sites are frequently changed by nursing females. Post-parturient females are normally very aggressive in response to any disturbance of their litter or the nest area. A female will pivot towards the source of any intrusion, erect her quills, puff loudly, emit a grating sound and roll her brow. She will exhibit a bucking response on close inspection, and attempt to bite. Cohabiting adults may also illicit a similar response, and males particularly tend to avoid nest chambers during the 10-14 days following parturition.
Unlike Spiny Hedgehog Tenrec Setifer setosus, in which the presence of a male or human disturbance invariably leads to cannibalism or rejection of neonates (Riordan, 1972), Echinops exhibits considerable social tolerance during parturition and the rearing of young. On certain occasions this tolerance has even extended to the successful rearing of more than one litter with several adults sharing a relatively small cage. Such a procedure is not recommended however, as it is doubtful whether individual mothers recognise their own young in the immediate post-partum period, and kidnapping and subsequent infant mortality is liable to occur (see Appendix). Instances of the cannibalism of infants have been recorded on only two occasions when a single pair shared a cage and in each case the male was responsible. We have no definite records of females cannibalising their own young.
Ideally therefore, pregnant females should still be isolated from all other animals during parturition and the rearing phase, though we have had many litters reared successfully with the sire remaining in the cage throughout this period. Moreover, even if the female is separated, neonates may also wander or be displaced from the nest and will be lost unless they are retrieved by the mother or can regain the refuge of the nest site by themselves. Retrieval of displaced infants is certainly characteristic of mothers during the early development of infants, but nursing females suddenly attracted from the nest box by live prey, may also carry suckling infants with them, and some of them may not be retrieved if they are scattering or wandering off. Daily checks on the progress of new litters are therefore carried out, if possible when the mother is away from the nest box. Infants that have been handled are not rejected however, and on one occasion two neonate infants whose mother died during parturition, were successfully fostered by another female in a separate cage. The latter had produced a litter of six at the same time and all eight infants were successfully reared.
Infant Growth and Development
In order to monitor the growth weights and development of individual animals within particular litters, infants were marked initially with indelible ink and later by clipping spines over different small areas of the dorsum. The latter works very satisfactorily, although it has to be repeated every few days during the first three or four weeks during rapid spine growth, but less frequently after that time. It is now also used for the long term marking of all adult individuals.
One of the most noticeable features of birth from Echinops is the marked disparity in birth weights between individuals in a single litter, see Tables I and II.
| Size and distribution of two live and six unborn young from a female which died during parturition on 6th July, 1978. |
| Condition |
Position in uterus |
Weight (gms) |
Length (mm) of head and body (nose to anterior margin of cloaca) |
Length (mm) of tail (Posterior margin of cloaca to tip of tail) |
|---|---|---|---|---|
| Live | left horn (?) | 7.2 | -- | -- |
| Live | left horn (?) | 7.9 | -- | -- |
| Dead | in vagina/cloaca | 6.2* | -- | -- |
| Dead | left horn | 9.0 | 52 | 7 |
| Dead | right horn | 9.4 | 58 | 7 |
| Dead | right horn | 6.7 | 50 | 7 |
| Dead | right horn | 6.7 | 53 | 5 |
| Dead | right horn | 8.1 | 54 | 6 |
| Weight range (gms) of neonates from three separate litters. |
| Litter 1 (Day 0) |
Litter 2 (Day 1) |
Litter 3 (Day 3) |
|
|---|---|---|---|
| 10.1 | 9.5 | 18.2 | |
| 8.0 | 6.2 | 18.3 | |
| 8.1 | 11.2 | 17.2 | |
| 7.4 | 9.1 | 17.2 | |
| 8.3 | 10.1 | -- | |
| 8.8 | 9.0 | -- | |
| 7.0 | 13.3 | -- | |
| 4.9 | 8.8 | -- | |
| 9.4 | -- | -- | |
| Ø= | 7.7 | 8.6 | 18.2 |
| n= | 9 | 8.6 | 4 |
In the first litter of Table II the two smallest infants (see Plate 2) failed to compete with their litter mates, and they both lost weight and died at five and seven days old respectively. The latter weighed only 3.1 gms. A third infant from the same litter did not grow as fast as its siblings throughout the rearing and weaning phase and eventually died at approximately eight weeks of age.
The small disparity in the size of neonates in the third litter of Table II is probably not related to the small size of the litter, since the overall incidence of runts has been quite high and runts have been recorded from small litters of three and four infants as well as in larger litters of eight or nine young. Litter size may, however, influence the future growth rates of young. Eisenberg and Gould (1966) have recorded an interval of as much as 30 hours between the birth of the first and last infants and they infer that the difference in time of birth favours the earliest born as they can suckle and gain strength even before a parturition is completed. In our experience parturitions are not normally spread over such a long period. Expectant mothers are generally checked every 24 hours and parturitions have invariably occurred during this period, usually during their diurnal phase. Moreover it is evident from Table I that the disparity in neonate weights is not directly related to the order of birth. The reason for the disparity itself is not known however, although it may, in part, be responsible for the relatively large percentage of females which have died during parturition, often from obstructed births -- see Appendix.
Echinops infants are born in an altricial state, with auditory meatus and eyes closed (Plate 1). They are nearly naked, though some spine growth is evident, but the degree of spinescence varies within a single litter. The young are capable of righting themselves if placed on their backs and can crawl forward for short distances. Some individuals (eg. four from a single litter of eight) were also able to roll themselves up when disturbed at this age. Grasping ability is well developed and clasping of fore and hind feet is characteristic. Individual variation in the colour and texture of spines is often marked at this age in litters from heterozygote morphs (see Plate 2) though these variations can be roughly distinguished into the two main categories of grey-skinned light-spined and pink-skinned brown-spined.
 Pigmy Hedgehog Tenrec Echinops telfairi at 36 days old.
Pigmy Hedgehog Tenrec Echinops telfairi at 36 days old.
Click on the photo or here to view a larger version.
Echinops infants mature very rapidly after birth and are functionally independent by 30-35 days of age. Eye opening occurs between the seventh and ninth day (Plate 3). Infants are more active by this time and are capable of orientation and seeking refuge themselves if displaced from the nest. By the tenth or ninth day they start to follow the mother when she leaves the nest and to wander and explore on their own. They may even abandon the nest site for much of the time and seek individual nesting places in crevices and logs in the immediate vicinity. All infants roll up and snort defensively when handled. Teeth are also erupt on the ninth or tenth day. The earliest observations of infants taking solid food have occurred at 13 days. By the fifteenth or sixteenth day all infants are taking solid food. They begin to forage by themselves and are capable of much faster movement as the ventrum is carried clear of the ground. They may also attempt to bite when handled. Mothers are far less protective towards their infants at this age, and although they will still defend the nest site, they no longer retrieve wandering young.
By eighteen or nineteen days of age, all infants will come to the food dish immediately it is introduced to the cage. The defensive 'bucking response' and grating of teeth is also evident on handling at this time, but the 'jump response' in catching live prey is not developed until about three weeks of age. By then, infants are beginning to compete for food and will try to steal food from litter mates and carry away food to consume it. Mothers may also steal food from young of this age and by the twenty-third or twenty-fourth days, feeding is characterised by all the animals rushing about with carried food, seeking privacy to consume it. From about two weeks of age females may nurse infants away from the nesting site, and this pattern continues as the infant develops and nursing will take place under any cover where the female or most infants may collect together. Nursing ceases at about 35 days of age (Plate 4).
Variations in the birth weights of infants are obviously exaggerated as weight is gained and this is also reflected in the overall size of infants. In view of the extremes that may be attained, total length does not seem to be a very dependable indicator of age as stated by Eisenberg and Gould (1970). This is particularly true when variations in litter size are also taken into consideration as larger litters grow relatively more slowly and exhibit more variation than small litters (see Figs. 6, 7 and 8).
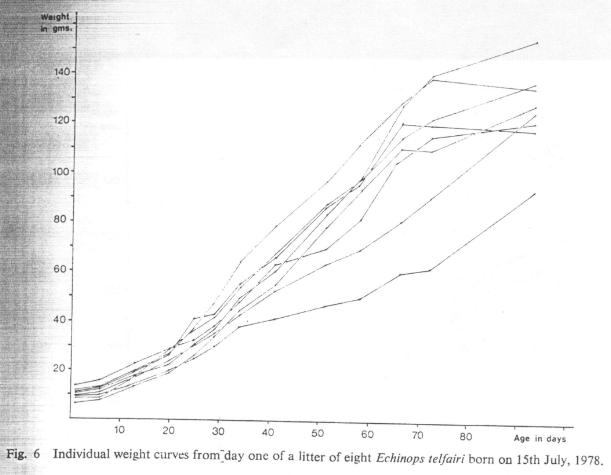 Fig. 6 Individual weight curves from day one of a litter of eight Echinops telfairi born on 15th July, 1978.
Fig. 6 Individual weight curves from day one of a litter of eight Echinops telfairi born on 15th July, 1978.
Click on the figure or here to view a larger version.
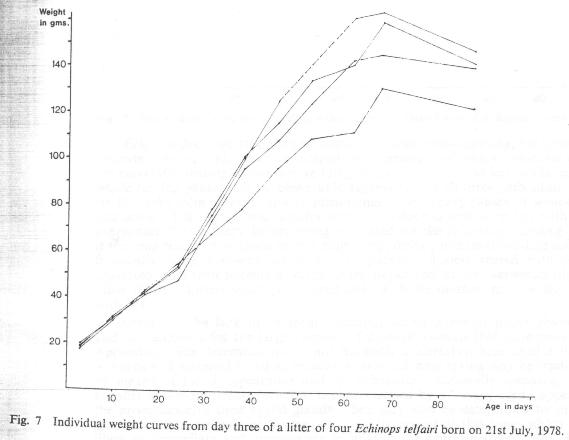 Fig. 7 Individual weight curves from day three of a litter of four Echinops telfairi born on 21st July, 1978.
Fig. 7 Individual weight curves from day three of a litter of four Echinops telfairi born on 21st July, 1978.
Click on the figure or here to view a larger version.
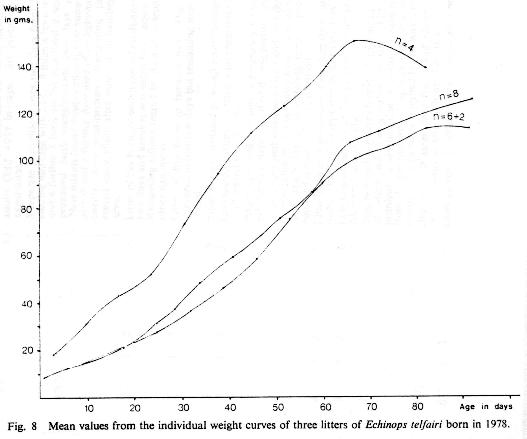 Fig. 8 Mean values from the individual weight curves of three litters of Echinops telfairi born in 1978.
Fig. 8 Mean values from the individual weight curves of three litters of Echinops telfairi born in 1978.
Click on the figure or here to view a larger version.
Echinops does not attain sexual maturity before over-wintering, but usually breeds the following season. However an interesting observation of male intolerance occurred after the successful rearing of five young (3 males, 2 females). The sire was left in the cage during the whole rearing phase with no observable aggression but all three (sub-adult) males died at 10-12 weeks from enteritis and/or pneumonia as secondary causes of wounds inflicted by this male. The two young females were not attacked and were left with the pair and overwintered with them, before being separated for the following breeding season. This is the only recorded instance of this happening, despite infants (including sub-adult males) frequently being overwintered with both parents. Litters reared with pairs are now separated from their parents at seven to ten weeks and left to overwinter together or with other litters. Litters which are reared only with the mother, may be left to overwinter with her.
Owing to the lack of external genitalia, sexing Echinops poses obvious difficulties and this accounts for the large number of unsexed animals that have been exported (see Appendix). Sex determination is not normally undertaken here until a litter has overwintered and assumed breeding condition, at which time sexing may be readily undertaken by means of facial appearance and on behaviour. Generally speaking, males have a broader and more robust face that is characterised by a 'baggy-eyed' appearance due to the prominance of their eyelid glands. Sex may also be determined by mixing unsexed specimens with animals of known sex (usually males) at this time, which will generally illicit an immediate and appropriate copulatory or aggressive response. In the case of males this response is invariably accompanied by copious eye gland secretion which in itself is a fairly reliable indicator of sex, though females may also occasionally exhibit the secretion (Poduschka, 1974). Eisenberg and Gould (1970) have successfully sexed several species of tenrec by the manual expression of the phallus or clitoris by manipulation of the cloacal area, but they found this procedure very difficult with Echinops owing to their rolling into a tight ball when disturbed. The technique has not yet been perfected here, though the animals rapidly become accustomed to handling and the rolling response will only be exhibited briefly unless the animals are roughly handled.
Appendix - Parturitions recorded at the Trust between 1967 and 1978.
| Litter No. |
Date of birth |
Gen. | Litter size | Reared | Comment | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mal | fem | unsex | tot | mal | fem | unsex | tot | ||||
| 1 | F1 | 2 | 1 | 1 | 4 | 2 | 1 | 1 | 4 | First parturitions from the two originally imported captive-bred yearling pairs. Both litters successfully reared with all four animals in the cage. | |
| 2 | F1 | 2 | 2 | 2 | 2 | ||||||
| 3 | F1 | 2 | 6 | 5 | 2 | 6 | 5 | Two parturitions comprising a total of eight infants born on consecutive days and all reared successfully with six adults (2 males 4 females) in cage. Six juveniles unsexed on exportation. | |||
| 4 | F1 | 3 | 3 | ||||||||
| 5 | F1 | 9 | 3 | 3 | 3 separate parturitions over two days from 3 of 4 females all sharing one cage with 2 males. 9 infants born to 2 females on 6th and a further 3 born on 7th to a third female who died from presumed birth complications two days post-partum. Nine infants found dead or missing over succeeding 20 day period and a further dam died of undetermined cause at twenty days post-partum. Remaining three infants reared successfully after removal for handrearing at that time. | ||||||
| 6 | |||||||||||
| 7 | F1 | 3 | |||||||||
| 8 | F1 | 1 | 2 | 2 | 5 | 1 | 2 | 3 | Litter of five born, of which one infant disappeared within two days and another was eaten by an adult at five days. Remaining infants reared by dam after separation from other specimens. | ||
| 9 | F1 | 1 | 3 | 1 | 5 | 1 | 3 | 4 | Litter of five born to isolated pair. One infant disappeared in first few days but remaining four successfully reared. | ||
| 10 | 1/2 F2 | 1 | 2 | 3 | 1 | 2 | 3 | Litter born to isolated pair - all reared successfully. | |||
| 11 | F2 | 1 | 2 | 3 | 1 | 2 | 3 | Late season parturition from a primiparous yearling dam - all reared successfully. | |||
| 12 | F2 | ? | Poorly recorded late season pregnancy from 3 year old primiparous dam who died during parturition - no p.m. | ||||||||
| 13 | F1 | 3 | 3 | Litter comprised one live (d.n.s.) and two dead foetuses (partly eaten by female cagemate) from an original imported dam which died after suffering from a prolapsed and ruptured uterus during parturition. | |||||||
| 14 | 1/2 F3 | 1 | 2 | 1 | 4 | 1 | 2 | 3 | Litter born to a primiparous yearling dam, sired by litter mate. 1 infant disappeared in first two days. | ||
| 15 | F3 | 1 | 2 | 3 | 1 | 2 | 3 | Late season parturition from a primiparous yearling dam. All reared successfully. | |||
| 16 | 1/2 F4 | 2 | Only two partially eaten foetuses found on cage floor, one of which was being eaten by sire. Litter born to primiparous yearling dam who died 3 days post-partum from chronic metritis. Staphylococcus was isolated from uterus and a partially resorbed foetus was found. | ||||||||
| 17 | F3 | 1 | Only a single large dead foetus and placenta found in nest. Foetus + afterbirth weighed 91/2 g. | ||||||||
| 18 | 1/2 F3 | 5 | 1 | 6 | 5 | 1 | 6 | Litter all reared successfully, sire removed from cage. | |||
| 19 | 1/2 F2 | 1 | 4 | 5 | 1 | 1 | Litter of two live and three dead young (one partially eaten) found in nest box with adult pair. One of the live infants disappeared after removal of sire, other successfully reared. | ||||
| 20 | F1 | 1 | 3 | 4 | 1 | 3 | 4 | Litter born to a recently imported pair which always produced litters in July in subsequent years. Litter reared with sire in cage. | |||
| 21 | 1/2 F4 | 1 | 1 | 1 | Unexpected parturition from recently imported female. Only second recorded singleton litter but since this infant was discovered some days after parturition, litter size is unreliable. The recent re-importation of the dam probably also explains the unusually late parturition. | ||||||
| 22 | F1 | 2 | 3 | 5 | 2 | 3 | 5 | Litter all reared successfully with sire in cage. | |||
| 23 | 1/2 F2 | 2 | 2 | 2 | 6 | 2 | 2 | 1 | 5 | Litter of six born to primiparous yearling dam, one infant disappeared in first week (no other adults present) others reared but one runt in litter died at 18 weeks when it weighed only 47 g despite treatment with growth hormones. | |
| 24 | F1 | 3 | 2 | 5 | 3 | 2 | 5 | Litter successfully reared to independence with sire present in cage, but all three males died of enteritis and/or pneumonia at 10-12 weeks following severe persecution by sire. | |||
| 25 | 1/2 F2 | 6 | 6 | 6 | 6 | Litter born to primiparous yearling female, all reared successfully with sire in cage. Unsexed on exportation. | |||||
| 26 | F2 | 2 | 2 | 2 | 2 | Litter of only two infants born to primiparous yearling female. Both reared successfully with sire in cage. Unsexed on exportation. | |||||
| 27 | F1 | 8 | 8 | 2 | 2 | Dam died during parturition, two infants born alive, third infant blocking birth passage. Further five full-term foetuses found dead in uterus. The two live infants fostered on to a second year female who gave birth to a primiparous litter of six on the same day. All eight infants were successfully reared but remain unsexed at the time of writing. | |||||
| 28 | 1/2 F3 | 6 | 6 | 6 | 6 | ||||||
| 29 | 1/2 F3 | 5 | 5 | Unborn full term litter, of which one large foetus obstructed birth passage resulting in death of the dam from toxaemia associated with obstructive dystocia. | |||||||
| 30 | 1/2 F2 | 8 | 8 | 8 | 8 | Large litter born to primiparous yearling dam - all reared but unsexed at time of writing. | |||||
| 31 | 1/2 F2 | 4 | 4 | 4 | 4 | Litter sired by 12 year old male, reared successfully with male in cage, but unsexed at time of writing. | |||||
| 32 | 1/2 F2 | 9 | 9 | 6 | 6 | Largest recorded litter. Born to a primiparous dam and reared with male in the cage (see fig.). 2 runts died within 2 days and one undeveloped infant died at 2 months - remainder exported at 3 months. | |||||
Acknowledgements
The authors gratefully acknowledge the assistance of Mr. N. Lindsay for the care and routine husbandry of the tenrecs under study and to Mr. Mark Vokins for his help with photography and assistance in other ways. We would also like to express our thanks to a number of other persons on the animal staff who assisted us at various times.
Finally we wish to acknowledge the assistance of the Wellcome Institute, Zoological Society of London and in particular to Dr. R. D. Martin.
References
Baker, F. J., Silverton, R. E., and Luckcock, E. D. (1966): Introduction to medical laboratory technology. Butterworths 4th edn. London, 656pp.
Eisenberg, J. F. and Gould, E. (1966): Notes on the biology of the Tenrecidae. J.Mamm., Vol. 47. 660-686. (Free Full Text)
Eisenberg, J. F. and Gould, E. (1967): The maintenance of tenrecoid insectivores in captivity. Int. Zoo Yb. 7. 194-196. (Free Full Text)
Eisenberg, J. F. and Gould, E. (1970): The Tenrecs. A study in Mammalian Behavior and Evolution. Smithsn. Contrib. Zool. 27, 1-137. (Free Full Text) (Alternative Free Full Text)
Eisenberg, J. F. and Muckenhirn, N. (1968): The reproduction and rearing of tenrecoid insectivores in captivity. Int. Zoo Yb. 8, 106-110. (Free Full Text)
Eisenberg, J. F. (1975): Tenrecs and Solenodons in captivity. Int. Zoo Yb. 15, 6-12. (Free Full Text)
Herter, K. (1962): Untersuchungen an lebenden Borstenigeln (Tenrecinae) 1: Über Temperaturregulierung und Aktivitätsrhythmik bei dem Igeltanrek Echinops telfairi Martin. Zoologische Beiträge, 7, 239-292.
Herter, K. (1963): Untersuchungen an lebenden Borstenigeln (Tenrecinae) 2: Über das Verhalten und die Lebensweise des Igeltanreks Echinops telfairi Martin, in Gefangenschaft. Zoologische Beiträge 8, 125-165.
Honegger, R. E. and Noth, W. (1966): Beobachtungen bei der Aufzucht von Igeltanreks Echinops telfairi Martin. Zoologische Beiträge, 12 (2), 191-218.
Mallinson, J. J. C. (1968): Conservation of Madagascan fauna in captivity - Tenrec collection. J.W.P.T. Fifth Annual Report 51-53. (Free Full Text)
Mallinson, J. J. C. (1972): Establishing mammal gestation periods at the Jersey zoological park. J.W.P.T. Ninth Annual Report 62-65.
Ormrod, S. A. (1967): Notes on tenrecoid. J.W.P.T. Fourth Annual Report 32-33. (Free Full Text)
Poduschka, W. (1974): Augendrüsensekretionen bei den Tenreciden Setifer setosus (Froriep 1806), Echinops telfairi (Martin 1838), Microgale dobsoni (Thomas 1918) und Microgale talazaci (Thomas 1918). Z. Tierpsychol., 35: 303-319.
Riordan, D. V. (1972): Reproduction in the spiny hedgehog tenrec Setifer setosus and the pigmy hedgehog tenrec Echinops telfairi. J.W.P.T. Ninth Annual Report 18-25.
Scaling, A. (1971): Progress report on the breeding of pigmy and spiny hedgehog tenrecs. J.W.P.T. Eighth Annual Report 12-14. (Free Full Text)
Weid, G. L. and Bibbo, M. (1971): Cytology of the female genital tract. In Pathway to conception. A. I. Sharman and C. C. Thomas (Ed). Springfield, Illinois 183-213.